Continual-Activation Theory of Dreaming
Jie Zhang
4418 Villa Paradiso Cres., Windsor, Ontario, Canada N9G 2L7
Abstract:
The theory presented in this paper hypothesizes that
both conscious and non-conscious subsidiary systems of working memory have to
be continually activated to maintain proper brain functioning. When the level of activation of either
subsidiary system descends to a given threshold, the continual-activation
mechanism in the brain will be triggered to generate a data stream from the
memory stores to flow through the subsidiary system in order to maintain the
activation level above this threshold. Dreaming (type II dream), as a conscious
experience, is only an epiphenomenon of brain continual-activation in the
conscious subsidiary system of working memory.
Key Words:
continual-activation theory, dreaming, memory, NREM, REM, working memory.
Introduction
Every person on earth dreams every night. But "Why do we dream?" or "What is the function of dreaming?" Although these questions have been the subject of a debate for centuries, the answers to these questions are still unknown.
In 1900, Sigmund Freud published his famous theory of dreams. He theorized that dreams involved complicated psychological processes that, when decoded, revealed the hidden desires of the dreamer. Freud's theory of dreams dominated sleep science in the first half of the 20th century until the 1950s, when Aserinsky and Kleitman (1953) discovered REM (rapid eye movement) sleep. Since then, many theories of dreaming have been proposed.
Allan Hobson and Robert McCarley (1977) presented a neurophysilogical model of dreaming, the Activation-Synthesis hypothesis. They hypothesized that dreaming resulted from the interpretation by the cortex of information concerning eye movements and activated brain stem motor pattern generators. They proposed that in REM sleep, the ascending cholinergic PGO (ponto-geniculo-occipital) waves stimulate higher midbrain and forebrain cortical structures, producing rapid eye movements. The activated forebrain then synthesizes the dream out of internally generated information. According to Hobson and McCarley "the forebrain may be making the best of a bad job in producing even a partially coherent dream imagery from the relatively noisy signals sent up to it from the brain stem." The model is based primarily on animal research of REM sleep, and assumed that the same structures that induce REM sleep also generate sensory information. The theory gives neither answer to the question why the brain should generate any images during REM sleep, nor explanation why this kind of vivid dream does not occur during waking, since the PGO waves occur at a fairly constant daily rate in the cat (14,000 + 3000 waves per day) (Jouvet, 1969), although it is markedly reduced in waking (Hobson, 2002). However, the activation-synthesis model, which has dominated the field of study of dreaming ever since, has been recently challenged by Mark Solms's neuropsychological – psychoanalytic model (Solms, 2000). Supported by the brain lesions study, Solms pointed out that dreaming and REM sleep are controlled by different brain mechanisms. That is, REM can occur without dreaming and dreaming can occur without REM.
Jie Zhang (2004) recently presented a new human memory model. By introducing a temporary memory stage into the memory process to bridge the gap between short-term memory and the long-term memory, Zhang proposed that during our waking time, the memory formed from the working memory is not saved directly into the long-term memory; instead it is saved into a temporary memory. The function of sleep is to process, encode and transfer the data from the temporary memory to the long-term memory. Zhang suggested that, besides the central executive (a control system that mediates attention and regulation of processes) (Baddeley & Hitch, 1974), the working memory could be divided into two groups of subsidiary systems: one for processing declarative (conscious) memory and one for processing procedural (non-conscious) memory. Corresponding to the type of memory been transferred, Zhang proposed that, sleep has two different stages: NREM (non rapid eye movement) sleep for processing the declarative memory, and REM sleep for processing the procedural memory. He further suggested that there are two types of dreams. The type I dream, a thought-like dream, is the result of the memory replay when the declarative memory is transferred from the temporary memory to the long-term memory during NREM sleep. The type II dream, a more dream-like dream, mainly occurs when the procedural memory is transferred from the temporary memory to the long-term memory during REM sleep. Based on this model, Zhang theorized that, although occurring often in REM sleep, the type II dream has no direct causality with the REM state.
Continual-Activation Theory
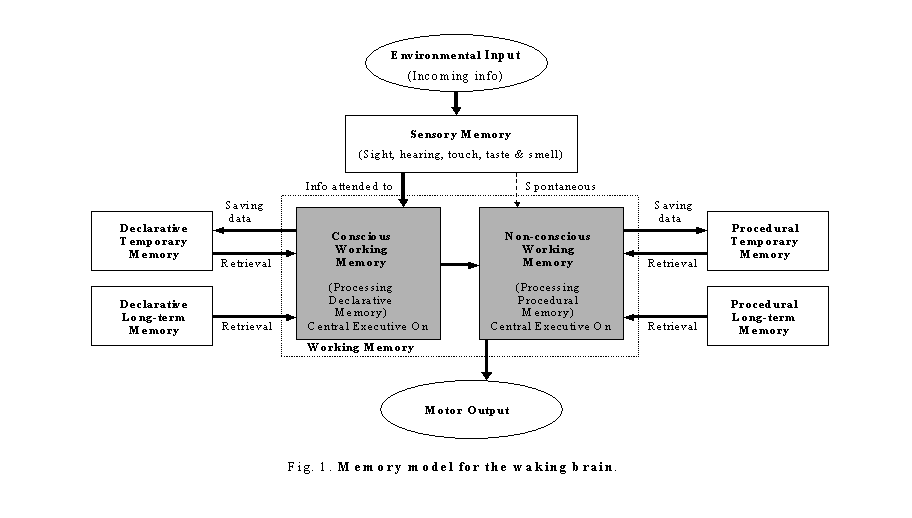
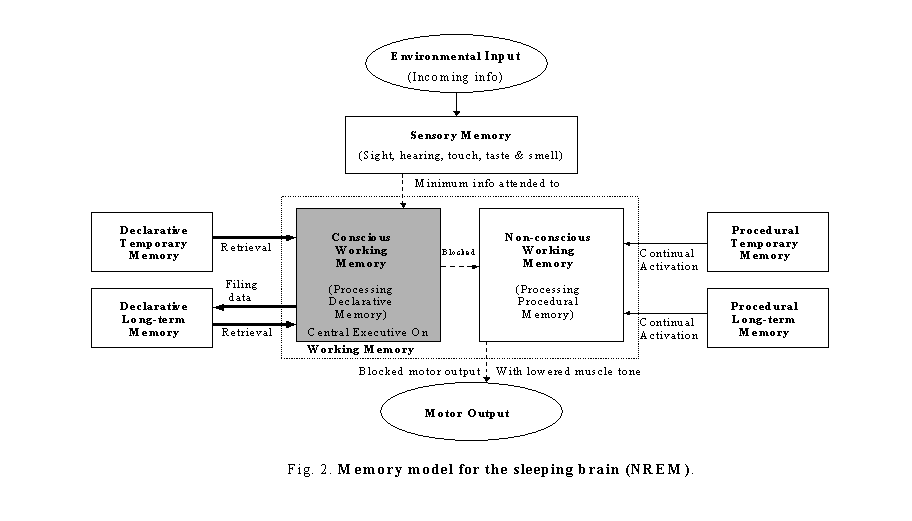
Continual-activation theory hypothesizes that both conscious and non-conscious subsidiary systems of working memory have to be continually activated to maintain proper brain functioning. When the level of activation of either subsidiary system descends to a certain threshold, the continual-activation mechanism in the brain will be triggered to generate a data stream from the memory stores to flow through the subsidiary system in order to maintain brain continual activation. To illustrate the continual-activation theory, a detailed Zhang memory model (Zhang, 2004) is given in figures 1 to 3.
During waking time, the sensory memory is continuously receiving information from all five senses. The information is first held in the sensory memory store. By attention, some of the information will then be passed to the conscious subsidiary system of working memory for processing. To process the incoming information, the conscious subsidiary system of working memory will retrieve related information from the declarative (conscious) temporary memory and or the declarative (conscious) long-term memory. The processed information will then be sent to the declarative temporary memory for rapid memory saving. In the meanwhile, the conscious subsidiary system of working memory will send a request to the non-conscious subsidiary system of working memory for a response output. To process this request, the non-conscious subsidiary system of working memory will also retrieve corresponding procedural memory from the procedural (non-conscious) temporary memory and or the procedural (non-conscious) long-term memory. The processed request will then be sent to the procedural temporary memory for rapid memory saving. At the same time, the processed request will also be passed to the motor output. The results of the motor output will then be monitored by five senses and fed back to the working memory for modifying. While awake, both the declarative and the procedural long-term memories are in retrieval-only mode. This process has been illustrated in figure 1. Here, the central executive will alternately pay attention to both subsidiary systems of working memory (illustrated by shaded symbols in the flow chart). During waking time, both the conscious and the non-conscious subsidiary systems of working memory are continually activated.
During NREM sleep period, the sensing rates of all the sensors are slowed and the arousal thresholds are increased. The central executive is now mainly engaged with the conscious subsidiary system of working memory for declarative memory processing/transferring. The conscious subsidiary system of working memory will retrieve data from the declarative temporary memory, compare it with related files previously saved in the declarative long-term memory, and delete any unwanted, duplicate or overlapping data. The information deemed relevant as new or updated, is encoded and transferred to the declarative long-term memory. While asleep, both declarative and procedural temporary memories are in retrieval-only mode. If awakened during this period time, the sleeper will have a thought-like mentation. This type of mentation has been defined as type I dream (Zhang, 2004). Sleep terrors may occur during this period of time when the brain is retrieving/processing/filling the most scary image memory data. Nevertheless, during this period of time, there is no attended data stream flowing through the non-conscious subsidiary system of working memory since the central executive (attention center) is busy with the conscious subsidiary system of working memory for memory processing. According to the continual-activation hypothesis, a continual-activation mechanism inside the brain will be triggered to automatically and randomly retrieve data from the procedural memory stores. This randomly retrieved data stream will flow through the non-conscious subsidiary system of working memory to maintain the brain continual activation (figure 2). Sleepers do not act out of this randomly retrieved procedural memory, because, by nature, their muscle tones are lowered. However, sometimes, for some reason, either the lowered muscle tone does not lower enough or the data stream is too strong, and the following sleep disorders might occur: sleep walking, sleep talking, tooth grinding and periodic limb movement disorder.
During the REM sleep period, the central executive is mainly engaged with the non-conscious subsidiary system of working memory for procedural memory processing/transferring. This time, the non-conscious subsidiary system of working memory will perform the data filling process similar to what happened in the conscious subsidiary system during NREM sleep. The so-called rapid eye movement (REM) is, in fact, a external manifestation of this filling process. Again, during this stage of sleep, the sleeper is prevented from acting out of this strong procedural memory data stream because the main muscle system is paralyzed. However, for some people this paralysis fails to occur during REM sleep, leading to REM sleep behavior disorder (RBD). As well, sometimes, the paralyzed muscle does not recover fast enough after waking, leading to sleep paralysis. During this period of time, there is no attended data stream flowing through the conscious subsidiary system of working memory. Similar to the case in NREM sleep, the continual-activation mechanism for the conscious subsidiary system of working memory will be initiated (figure 3). This mechanism will randomly retrieve declarative memory data from the declarative memory stores to feed to the conscious subsidiary system of working memory in order to meet the minimum level required for brain activation. The easiest, most accessible information, such as day residue, will be more salient for retrieval. Since the data stream comes from random activation, the sleeper should have a mentation similar to the type I dream during this time. One thing that needs to be pointed out is that not every type I dream can be recalled: the dream without visual content is surely more difficult to recall. However, as the randomly retrieved memories flow through the conscious subsidiary system of working memory, the brain will attempt to make sense and interpret them. With the involvement of the brain association (associative thinking) system (believed to be mainly looped through the area of the frontal lobes) and the emotion system, the vivid dreaming event (type II dream) will start shortly after. The level of brain activation in the conscious subsidiary system of working memory will then be increased to a higher level. Based on the dreamer's thinking, the association system will associatively retrieve memories from the memory stores and quickly turn them into pictures or realistic-like events. This may, sometimes, bring dreamers a feeling of their thoughts being broadcast, as some of their thoughts (judgment, decision, etc.) end within the next dream scene in someway. Although the data randomly retrieved by the continual-activation mechanism will be interpreted by the dreaming brain and synthesized into a dream plot, the type II dream plot is strongly influenced by the dreamer's own thinking. From this point of view, it is easy to understand that a person with brain damage to the frontal lobes (association system), such as the lobotomy patients, or to the "visual-spatial sketch-pad" (spatial imagery loop) (Baddeley & Hitch, 1974) may not have a vivid dream-like dream (type II dream), although the continual-activation mechanism is still functioning.
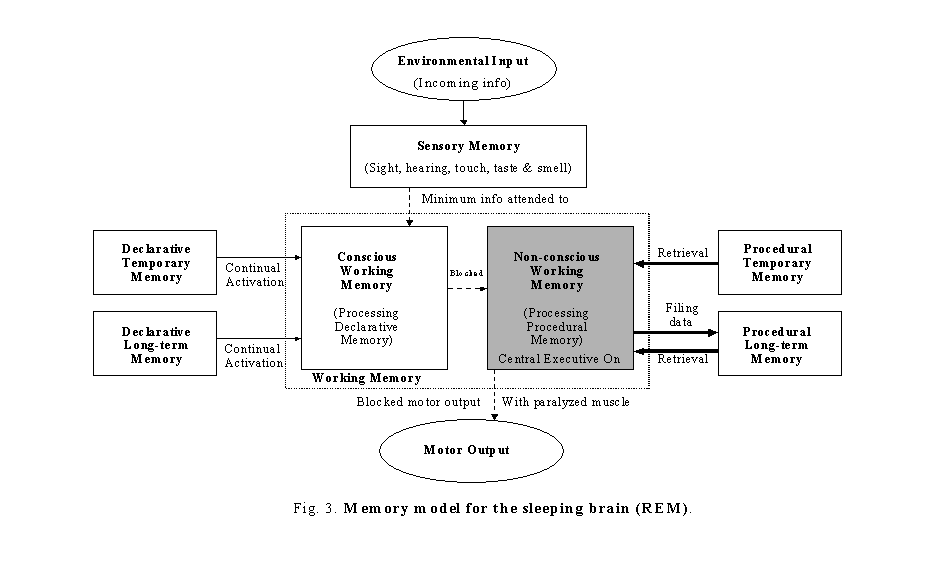
As we shall now see, the continual-activation theory of dreaming hypothesizes that the type II dream is a result of involvement of the conscious subsidiary systems of working memory with this continual-activation mechanism. The theory is in agreement with Hobson and McCarley's Activation-Synthesis hypothesis in the aspect of that dreaming (type II dream) is based on physiological mechanisms not psychic forces, and is the results of brain activation and synthesis. However, the continual-activation theory differs with the activation-synthesis hypothesis in one main ground. The continual-activation theory proposes that REM sleep and dreaming have different physiological mechanisms and serve different functional purposes. Dreaming can occur without REM and REM can occur without dreaming which support Mark Solms's finding (Solms, 2000).
Conclusion
Besides the central executive, the working memory could be divided into two subsidiary systems: the conscious and the non-conscious subsidiary systems. Both subsidiary systems have to be continually activated through their life times. It is very much like the brain has "hearts" of its own. When the flow of data stream in either subsidiary system drops to a given threshold, a continual-activation mechanism will be triggered to randomly retrieve memory data from the memory stores to maintain brain activation. Dreaming, as a conscious experience, is only a result of the involvement of conscious subsidiary system of the brain with this continual-activation data stream.
Acknowledgements: The author would like to thank Jeremy Tebbens and Bill Boyle for valuable discussions on this manuscript.
References
- Aserinsky, E. & Kleitman, N. (1953) Regularly occurring periods of eye motility and concurrent phenomena during sleep. Science. 118, 273-274.
- Baddeley, A.D. and Hitch, G.J. (1974) Working memory. The psychology of learning and motivation. Academic press. New York, 47-89.
- Hobson, J.A & McCarley, R. (1977) The brain as a dream state generator: An activation-synthesis hypothesis of the dream process. American Journal of Psychiatry 134, 1335-1348.
- Hobson, J.A. (2002) Dreaming-an introduction to the science of sleep. Oxford University Press. New York.
- Jouvet, M. (1969) Biogenic amines and the states of sleep, Science 163(862), 32-41.
- Solms M. (2000) Dreaming and REM sleep are controlled by different brain mechanisms. Behavioral and Brain Sciences 23(6), 793-1121.
- Zhang, J. (2004) Memory process and the function of sleep. Journal of Theoretics Vol.6-6.